Florística y estructura del componente Arbustivo y Arbóreo en el Parque Estatal de Itapeva, RS
Régis Cardoso Silva1*, Robson Santos1, Dilton Pacheco1, Rafael Martins1, Vanilde Citadini-Zanette1 Ronaldo Josué Faller 2
1Universidade do Extremo Sul Catarinense - UNESC
2 Universidad Federal de Rio Grande do Sul – UFRGS
*Autor para correspondência E-mail: registrs37@gmail.com
Recebido: 15.10. 2023 Aceito: 18.05. 2024
RESUMEN: Con el objetivo de aumentar nuestro conocimiento sobre los bosques arenosos de restinga en Rio Grande do Sul, realizamos un estudio florístico y fitosociológico del componente arbustivo de un capón de restinga en el Parque Estadual de Itapeva (PEVA). Este estudio utilizó el método de parcelas, incluyendo todos los individuos con un DAS (diámetro a la altura del suelo) mínimo de 2,5 cm y totalizando un área de 0,5 ha. La composición florística dio como resultado una riqueza total de 81 especies. La densidad total de arbustos en el capão fue de 4.240 ind/ha. La familia con mayor riqueza específica fue Myrtaceae y entre las especies con mayor valor de importancia estuvieron Myrsine umbellata Mart. y Sorocea bonplandii (Baill.) W. C. Burger. En regeneración natural, las 10 primeras especies representaron el 54% de la regeneración natural total, con tasas que oscilaron entre el 13,5% y el 0,2%. La elevada riqueza observada demostró la contribución de la "Porta de Torres" como importante corredor migratorio de especies atlánticas en la vegetación de Rio Grande do Sul, añadiendo nuevas informaciones sobre la florística y estructura comunitaria de la Restinga de Rio Grande do Sul. Los resultados presentados contribuirán a nuevos estudios sobre la dinámica del PEVA y a programas de gestión, preservación y recuperación ambiental.
Palabras-clave: Restinga; Estructura fitosociológica; Mata Atlántica.
Floristics and structure of the Shrub-Tree component of the Itapeva state park, RS
ABSTRACT: To enlarge the knowledge on sandy Restinga forests in Rio Grande do Sul, it was carried out a shrub-tree component phytosociological and floristic studies of a Restinga forest from Parque Estadual de Itapeva (PEVA). In this study, it was used the the plot method, including all the individuals with DAS (soil height diameter) minimum of 2.5 cm and totaling an area of 0.5 ha. The floristic composition resulted in a total richness of 81 species. The shrub-tree total density in the forest was 4,240 individulas/ha. The family with the highest specific richness was Myrtaceae and, among the species with the highest importance value are Myrsine umbellata Mart. and Sorocea bonplandii (Baill.) W. C. Burger. In the natural regeneration the 10 first species accumulated 54% of the total natural regeneration, with rates from 13.5% to 0.2%. The high observed richness showed the contribution of "Porta de Torres" as an important Atlantic species migratory corridor in the vegetation in Rio Grande do Sul, adding new information on Restinga community structure and floristic in Rio Grande do Sul. The results will contribute to further studies on PEVA dynamic and to environmental reclamation, preservation and handling programs.
Keywords: Sandy dune vegetation; phytosociological structure; Atlantic Forest.
Florística e estrutura do componente Arbustivo-Arbóreo do Parque Estadual de Itapeva, RS
RESUMO: Com o objetivo de ampliar nosso conhecimento sobre as florestas arenosas de restinga no Rio Grande do Sul, realizamos um estudo florístico e fitossociológico do componente arbustivo de um capão de restinga no Parque Estadual de Itapeva (PEVA). Este estudo utilizou o método de parcelas, incluindo todos os indivíduos com um DAS mínimo (diâmetro à altura do solo) de 2,5 cm e totalizando uma área de 0,5 ha. A composição florística resultou em uma riqueza total de 81 espécies. A densidade total de arbustos no capão foi de 4.240 ind/ha. A família com maior riqueza específica foi Myrtaceae e entre as espécies com maior valor de importância estão Myrsine umbellata Mart e Sorocea bonplandii (Baill.) W. C. Burger. Na regeneração natural, as 10 principais espécies representaram 54% da regeneração natural total, com taxas variando de 13,5% a 0,2%. A alta riqueza observada demonstrou a contribuição da Porta de Torres como importante corredor migratório de espécies atlânticas na vegetação gaúcha, agregando novas informações sobre a florística e estrutura da comunidade da Restinga do Rio Grande do Sul. Os resultados apresentados contribuirão para novos estudos sobre a dinâmica do PEVA e para programas de gestão, preservação e recuperação ambiental.
Palavras-chave: Restinga; Estrutura fitossociológica; Mata Atlântica.
INTRODUCIÓN
En general, las restingas presentan especies procedentes de otros ecosistemas que han colonizado estos ambientes debido a la variedad de condiciones físicas que allí se dan (Araújo; Lacerda, 1987; Freire, 1990), pero exhiben variaciones fisonómicas debido a las diferentes condiciones de su ambiente de origen (Assumpção; Nascimento, 2000; Magnago et al., 2010). Las especies vegetales presentes en estas comunidades tienen características que les permiten colonizar suelos pobres, arenosos, encharcados o muy salinos, y ambientes con vientos constantes (Waechter, 1985).
La vegetación de restinga desempeña muchas funciones en el ecosistema donde se encuentra. Algunos autores asocian esta vegetación a la estabilización del sustrato en estos ambientes, como protección de la acción de los vientos, lo cual se considera un modificador muy importante del paisaje (Lindeman, 1906; Rambo, 1954; Assumpção; Nascimento, 2000). Además, contribuye al mantenimiento del drenaje natural y a la preservación de la fauna endémica y migratoria (Falkenberg, 1999; Rocha et al., 2005).
En el contexto de Rio Grande do Sul, las dos principales rutas migratorias de las especies vegetales tropicales se encontraban en el lado este, sirviendo de corredor para las especies tropicales atlánticas, y en el lado noroeste, constituido por especies típicamente estacionales (Rambo, 1950). Estas especies tropicales migraban hacia el estado a través de la "Porta de Torres", la estrecha llanura entre la Meseta Sur y el Océano Atlántico, descrita por Rambo (1950) como una importante ruta migratoria para las especies tropicales del sudeste de Brasil.
El municipio de Torres es una importante puerta de entrada para la vegetación de Rio Grande do Sul, según la descripción de Rambo (1950). Este corredor incluye el Parque Estatal de Itapeva (PEVA), que viene sufriendo las consecuencias de la actividad humana. Su principal impacto incluye la introducción de especies animales y vegetales exóticas, la ocupación humana y la presión agrícola, así como el crecimiento de la ciudad de Torres dentro de los límites del Parque.
Para comprender mejor las zonas de bancos de arena, es necesario describir la vegetación, lo que puede hacerse básicamente mediante métodos florísticos y estructurales (Kent; Coker, 1992). Así, la enumeración de la flora y la caracterización de la estructura contribuyen a además de diferenciar sus fisonomías (Silva; Britez, 2005; Magnago et al., 2011), proporcionando más conocimiento sobre las restingas del sur de Brasil.
Teniendo en cuenta estas cuestiones, el objetivo de este estudio fue conocer la estructura y composición florística de la restinga arbustiva y arbórea del Parque Estadual de Itapeva, en Torres, Rio Grande do Sul, así como relacionar la composición florística con diferentes estudios realizados en el sur de la Mata Atlántica brasileña.
1. MATERIAL Y MÉTODOS
- Localización de la zona
El estudio se realizó en el Parque Estadual de Itapeva (PEVA), que abarca un área total de 1.000 ha, localizado en el municipio de Torres, en el litoral norte de Rio Grande do Sul (29º 20' S y 49º 45' O) (Figuras 1 y 2). El clima dela región, según la clasificación de Köppen, es del tipo Cfa, mesotérmico húmedo, con inviernos suaves y veranos calurosos. La precipitación media anual es de 1.535 mm y la temperatura media anual es de 19,3 °C (INMET, 2009). Este parque está ubicado aproximadamente a 200 km de Porto Alegre, la capital del estado.
- Metodología
Se asignaron un total de 100 parcelas de 5 m x 5 m, incluyeondo todos los individuos con un diámetro mínimo del tallo al nivel del suelo de 2,5 cm, totalizando en una superficie de muestreo de 0,25 ha. Este procedimiento se adoptó debido a que la zona estudiada es un cordón arenoso ondulado, con dunas móviles próximas a la vegetación estudiada, siguiendo las recomendaciones de Felfili et al. (2011). Los datos y las muestras botánicas se recogieron entre diciembre de 2008 y mayo de 2009, teniendo en cuenta únicamente a arbustos y árboles. Los descriptores fitosociológicos de frecuencia, densidad, dominancia y valor de importancia se calcularon según Mueller-Dombois y Ellenberg (1974). El material fértil colectado fue depositado en el Herbario del Padre Dr. Raulino Reitz (CRI) de la Universidad del Extremo Sur de Santa Catarina (UNESC). La actualización taxonómica de las especies y sus autores siguió las propuestas de Forzza et al. (2010).
Para cada especie de arbusto, se proporcionó información sobre el grupo ecológico y estrategia de dispersión. Los grupos ecológicos fueron definidos siguiendo la propuesta de Ferretti et al. (1995), mientras que las estrategias de dispersión se basaron en los principios de Van der Pijl (1972).
Para el estudio de la regeneración natural, se establecieron las siguientes clases según Volpato (1994): clase 1, formada por individuos con una altura igual o superior a 20 cm e inferior a 50 cm; y clase 2, formada por individuos con una altura igual o superior a 50 cm y un DAP inferior a 2,5 cm. El muestreo se realizó en 100 parcelas de 1 m x 1 m y 2 m x 2 m, respectivamente, para cada clase de altura. Se calculó el Potencial de Regeneración Natural (PRN) para las dos clases consideradas, que es un parámetro sintético derivado de la abundancia (densidad) y distribución (frecuencia) de las diferentes clases de tamaño. También se estimó el Potencial de Regeneración Natural Total (TNR) de la especie, que evalúa la regeneración total de la especie a través de la suma de las clases de tamaño consideradas, según Finol (1971), modificada por Volpato (1994).
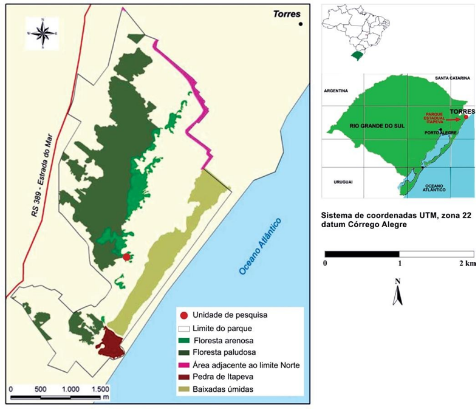
Figura 1 - Localización y clasificación de las fisonomías encontradas en el Parque Estadual de Itapeva, municipio de Torres, Rio Grande do Sul. Fuente: SEMA (2006), modificado.
Figure 1 - Location and classification of physiognomies found in the Itapeva State Park, municipality of Torres, Rio Grande do Sul. Source: SEMA (2006), as amended.
Figura 1 - Localização e classificação das fisionomias encontradas no Parque Estadual de Itapeva, município de Torres, Rio Grande do Sul. Fonte: SEMA (2006), conforme alterada.
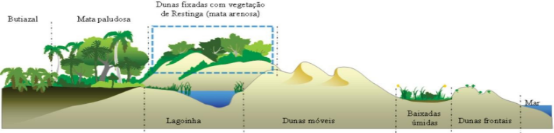
Figura 2 - Perfil esquemático de las fitofisonomías encontradas en el Parque Estatal de Itapeva, en el municipio de Torres, RS, con el perfil del bosque arenoso estudiado resaltado.
Figure 2 - Schematic profile of the phytophynonomies found in the Itapeva State Park, in the municipality of Torres, RS, with the profile of the sandy forest studied highlighted.
Figura 2 - Perfil esquemático de las fitofisonomías encontradas en el Parque Estatal de Itapeva, en el municipio de Torres, RS, con el perfil del bosque arenoso estudiado resaltado.
Para relacionar el área de estudio y su patrón de distribución con diferentes estudios realizados en bosques de restinga y áreas de bosque denso ombrofilo, se construyeron matrices de datos binarios (0 para ausencia y 1 para presencia). Para ello se consultó a: Citadini-Zanette (1995) FODS1; Citadini- Zanette et al. (2001) R E S T 1; Dillenburg et al. (1992) REST2; Dorneles y Waechter (2004) REST3; Hentschel (2008) REST5; Jarenkow (1994) FODM1; Martins (2005) FODS2; Martins (2010) FODM2; Moraes y Mondin (2001) REST6; Negrelle (2006) FOTB1; este estudio REST4; Rossoni y Baptista (1995) REST7; Scherer et al. (2005) REST8; y Silva (1990) REST9. Posteriormente, estas matrices fueron analizadas utilizando el análisis de correspondencia (AC), que permite verificar la ocurrencia de gradientes en el ordenamiento de las muestras (Kent y Coker, 1992), dispuestas en un diagrama bidimensional.
2. RESULTADOS
Se registró un total de 81 especies distribuidas en 55 géneros y 30 familias botánicas. Se tomaron muestras de 1.060 individuos, con una densidad total de 4.240 individuos. ha-1 (Tabla 1). De las familias registradas, 13 (16%) estaban representadas por una sola especie, tres (7%) por dos especies y ocho (30%) por tres especies. Myrtaceae presentó la mayor riqueza específica, com 16 especies (20%), de las cuales siete pertenecen al género Myrcia y cuatro al género Eugenia. Le siguieron las Lauraceae con 10 especies (12%) y las Anacardiaceae, Euphorbiaceae y Rubiaceae con cuatro especies cada una (15%). Estas cinco familias representaban el 47% de las especies muestreadas.
Se identificaron un total de 46 especies arbustivas y arbóreas que colonizan zonas de bosque arenoso, que no se mencionaban en el Plan de Gestión del Parque para este entorno, así como otras 14 que no se habían mencionado para el PEVA. Se trata de Chrysophyllum viride, Cryptocarya moschata, Diospyros inconstans, Erythroxylum deciduum, Faramea montevidensis, Ficus cestrifolia, Myrceugenia cf. glaucescens, Myrcia campestris, M. tijucensis, Nectandra lanceolata, Ouratea salicifolia, Piper gaudichaudianum, Randia ferox y Zanthoxylum astrigerum.
En términos de densidad, Sorocea bonplandii destacó con el mayor número de individuos (372 individuos. ha-1). En términos de frecuencia, Myrsine umbellata tuvo la mayor ocurrencia, estando presente en 44 de las 100 unidades de muestreo. El valor obtenido para el área basal total fue de 41,6 m². ha-¹, siendo la especie con mayor dominancia y contribución a este parámetro M. umbellata, con 4,64 m².ha-¹.
Los valores de diámetro (DAS) oscilaron entre 2,5 cm y 63,7 cm. Aproximadamente el 44,8% del total tenían entre 2,5 y 4,9 cm; el 53,3% entre 5,0 y 29,9 cm; y sólo el 2% tenían tallos iguales o superiores a 30,0 cm. El mayor valor diametral lo obtuvo Ficus cestrifolia (63,7 cm), seguido de individuos de Chrysophyllum inornatum y Coussapoa microcarpa, con diámetros de 60,8 cm y 59,5 cm, respectivamente.
Las especies con mayor valor de importancia (VI) fueron Myrsine umbellata, Sorocea bonplandii, Eugenia brevistyla, Chrysophyllum inornatum, Nectandra lanceolata, N. oppositifolia, Myrcia glabra, Aiouea saligna, Coussapoa microcarpa y Cupania vernalis. Estas 10 especies representaban el 42,6% de los individuos muestreados.
La altura de los individuos registrados en este estudio oscilaba entre 1 y 13 metros, con una altura media de 4,4 metros. Las especies encontradas en el estrato inferior de la zona de estudio fueron Sorocea bonplandii, Gymnanthes concolor, vernalis y Eugenia bacopari. Aproximadamente el 55% de los individuos muestreados se concentraban en el estrato inferior. En el estrato medio, Eugenia brevistyla, Myrsine umbellata, Sorocea bonplandii, Myrcia glabra y Chrysophyllum inornatum fueron las más representativas. Entre las especies concentradas en el estrato superior, destacaron Maytenus sp., Myrcia pubipetala y Ficus luschnathiana, con 13, 12 y 11 m, respectivamente.
Tabla 1 - Parámetros estructurales para especies arbustivas en 0,25 ha de un bosque de restinga arenosa en el Parque Estatal de Itapeva, Torres, RS. Para individuos con DAP > 2,5 cm. Donde: H- altura máxima (m); FA- frecuencia absoluta (%); DoA- dominancia absoluta (m².ha- ¹); DA- densidad absoluta (individuos.ha- ¹); VI- valor de importancia; Eco - grupo ecológico, donde: P- pionero, Si- secundario temprano, St- secundario tardío y C- clímax; SD- síndrome de dispersión, donde: An- anemocoria, Au- autocoria y Z- zoocoria.
Table 1 - Structural parameters of shrub-tree species in 0.25ha in a Restinga forest in the State Park of Itapeva, Torres, RS, for individuals with DAS > 2.55cm. Where: H- maximum height (m); FA- absolute frequency (%), DoA- absolute dominance (m².ha-¹), DA- absolute density (individuals.ha-¹), VI- importance value, ECO- ecological group, where, P- pioneer, Si- initial secondary, St- late secondary and C- climax; SD- dispersion syndrome, where, An- anemochoric, Au- autochoric and Z- zoochoric.
Tabela 1 - Parâmetros estruturais para espécies arbustivas em 0,25 ha de floresta arenosa de restinga no Parque Estadual de Itapeva, Torres, RS, para indivíduos com DAP > 2,5 cm. Onde: H- altura máxima (m), AF- frequência absoluta (%), DoA- dominância absoluta (m². ha- ¹), DA- densidade absoluta (indivíduos.ha-¹), VI- valor de importância, ECO - grupo ecoecológico, onde, P- pioneiro; Sim- secundário precoce; St- secundários tardios e C-clímax; SD- síndrome de dispersão, onde, An- anemocoria, Au- autocoria e Z- zoocoria.
Especie | H | FA | DoA | DA | VI | Eco | SD |
Myrsine u m b e l l a t a Mart. | 13,0 | 44 | 4,639 | 260 | 7,89 | Sí | Z |
Sorocea bonplandii (Baill.) W.C.Burger et al. | 6,0 | 41 | 1,162 | 372 | 5,84 | San | Z |
Eugenia brevistyla D.Legrand | 9,0 | 33 | 2,967 | 208 | 5,61 | P | Z |
Chrysophyllum inornatum Mart. | 11,0 | 30 | 2,188 | 248 | 5,15 | C | Z |
Nectandra lanceolata Nees et Mart. | 11,0 | 19 | 2,739 | 116 | 4,02 | C | Z |
Nectandra oppositifolia Nees et Mart. | 12,0 | 16 | 2,924 | 84 | 3,78 | San | Z |
Myrcia glabra (O. Berg) D. Legrand | 9,0 | 24 | 1,174 | 168 | 3,42 | San | Z |
Aiouea saligna Meisn. | 9,0 | 26 | 1,480 | 124 | 3,42 | Sí | Z |
Coussapoa microcarpa (Schott) Rizzini | 11,0 | 11 | 2,706 | 64 | 3,20 | San | Z |
Cupania vernalis Cambess. | 7,5 | 27 | 0,756 | 164 | 3,20 | San | Z |
Alchornea triplinervia (Spreng.) Müll. Arg. | 12,0 | 15 | 2,037 | 80 | 2,99 | Sí | Z |
Ficus cestrifolia Schott | 7,5 | 11 | 2,582 | 44 | 2,95 | P | Z |
Myrcia pubipetala Miq. | 12,0 | 17 | 1,654 | 100 | 2,93 | San | Z |
Gymnanthes concolor Spreng. | 6,0 | 17 | 0,259 | 164 | 2,32 | San | Au |
Solanum pseudoquina A. St.-Hil. | 12,0 | 14 | 1,187 | 80 | 2,26 | P | Z |
Marlierea eugeniopsoides (Kausel & D. Legrand) D. Legrand | 10,0 | 12 | 0,965 | 104 | 2,17 | C | Z |
Psidium cattleianum Sabine | 8,0 | 19 | 0,642 | 88 | 2,12 | San | Z |
Eugenia bacopari C. D. Legrand | 6,5 | 15 | 0,345 | 128 | 2,01 | P | Z |
Ocotea pulchella (Nees) Mez | 11,0 | 14 | 0,833 | 68 | 1,88 | C | Z |
Matayba guianensis Aubl. | 10,0 | 13 | 0,611 | 76 | 1,72 | San | Z |
Maytenus sp. | 13,0 | 14 | 0,194 | 104 | 1,65 | San | Z |
Mollinedia schottiana (Spreng.) Perkins | 5,0 | 13 | 0,227 | 100 | 1,60 | C | Z |
Myrcia brasiliensis Kiaersk. | 7,0 | 13 | 0,366 | 60 | 1,39 | San | Z |
Zanthoxylum rhoifolium Lam. | 9,0 | 11 | 0,575 | 44 | 1,34 | Sí | Au |
Guapira opposita (Vell.) Reitz | 12,0 | 11 | 0,381 | 60 | 1,31 | Sí | Z |
Cabralea canjerana (Vell.) Mart. | 7,5 | 12 | 0,257 | 60 | 1,26 | San | Z |
Esenbeckia grandiflora Mart. | 5,0 | 10 | 0,070 | 88 | 1,23 | San | Au |
Allophylus edulis (A. St.-Hil., Cambess. et A. Juss.) Radlk. | 7,0 | 11 | 0,292 | 48 | 1,14 | Sí | Z |
Mollinedia sp. | 4,0 | 8 | 0,096 | 60 | 0,94 | C | Z |
Endlicheria paniculata (Spreng.) J. F. Macbr. | 9,0 | 6 | 0,246 | 52 | 0,90 | San | Z |
Symphyopappus casarettoi Robinson | 6,0 | 8 | 0,217 | 40 | 0,87 | P | En |
Schinus polygamus (Cav.) Cabrera | 8,0 | 4 | 0,575 | 28 | 0,87 | P | Z |
Schinus terebinthifolius Raddi | 4,0 | 7 | 0,245 | 40 | 0,85 | P | Z |
Sapium glandulosum (L.) Morong | 6,0 | 8 | 0,102 | 48 | 0,85 | P | Z |
Myrsine lorentziana (Mez) Arechav. | 7,0 | 8 | 0,169 | 40 | 0,84 | Sí | Z |
Ficus luschnathiana (Miq.) Miq. | 11,0 | 3 | 0,623 | 12 | 0,74 | San | Z |
Blepharocalyx salicifolius (Kunth) O. Berg | 6,0 | 7 | 0,123 | 36 | 0,72 | P | Z |
Erythroxylum cuspidifolium Mart. | 5,5 | 7 | 0,063 | 40 | 0,70 | C | Z |
Posoqueria latifolia (Rudge) Roem. et Schut. | 4,0 | 8 | 0,051 | 32 | 0,68 | Sí | Z |
Garcinia gardneriana (Planch. et Triana) Zappi | 6,0 | 6 | 0,072 | 40 | 0,66 | San | Z |
Cryptocarya moschata Nees et Mart. ex Nees | 10,0 | 3 | 0,424 | 16 | 0,61 | C | Z |
Erythroxylum deciduum A. St. Hil. | 7,0 | 5 | 0,261 | 20 | 0,61 | P | Z |
Ouratea parviflora (DC.) Baill. | 4,5 | 7 | 0,029 | 28 | 0,58 | Sí | Z |
Myrsine coriacea (Sw.) R. Br. ex Roem et Schult. | 6,0 | 5 | 0,202 | 20 | 0,56 | P | Z |
Eugenia schuechiana O. Berg | 5,0 | 6 | 0,091 | 24 | 0,55 | C | Z |
Faramea montevidensis (Cham. et Schltdl.) DC. | 6,0 | 4 | 0,082 | 32 | 0,51 | C | Z |
Casearia sylvestris Sw. | 6,0 | 5 | 0,089 | 24 | 0,50 | Sí | Z |
Persea willdenovii Kosterm. | 10,0 | 4 | 0,167 | 20 | 0,48 | San | Z |
Randia ferox (Cham. et Schltdl.) DC. | 5,0 | 5 | 0,037 | 24 | 0,46 | Sí | Z |
Lithraea brasiliensis Marchand | 4,0 | 4 | 0,078 | 24 | 0,44 | P | Z |
Hirtella hebeclada Moric. ex DC. | 4,5 | 3 | 0,114 | 20 | 0,39 | San | Z |
Piper gaudichaudianum Kunth | 4,5 | 4 | 0,036 | 20 | 0,38 | Sí | Z |
Prunus myrtifolia (L.) Urb. | 5,0 | 4 | 0,070 | 16 | 0,38 | Sí | Z |
Trichilia lepidota Mart. | 8,0 | 3 | 0,076 | 12 | 0,30 | C | Z |
Ocotea puberula (Reich.) Nees | 8,0 | 1 | 0,258 | 4 | 0,29 | C | Z |
Miconia sellowiana Naudin | 7,0 | 3 | 0,050 | 12 | 0,28 | Sí | Z |
Persea venosa Nees | 5,5 | 2 | 0,111 | 12 | 0,28 | San | Z |
Ilex theezans Mart. ex Reissek | 6,5 | 2 | 0,060 | 12 | 0,24 | P | Z |
Myrceugenia cf. glaucescens (Cambess.) D. Legrand & Kausel | 5,0 | 1 | 0,199 | 4 | 0,24 | San | Z |
Myrcia multiflora (Lam.) DC. | 6,0 | 2 | 0,050 | 12 | 0,23 | C | Z |
Casearia decandra Jacq. | 3,5 | 1 | 0,055 | 16 | 0,22 | P | Z |
Myrcia palustris DC. | 2,0 | 2 | 0,022 | 8 | 0,18 | Sí | Z |
Tetrorchidium rubrivenium Poepp. Myrcia campestris DC | 3,0 | 2 | 0,017 | 8 | 0,17 | Sí | Z |
Chrysophyllum viride Mart. et Eichler | 3,0 | 1 | 0,017 | 8 | 0,13 | C | Z |
Schinus sp. | 3,0 | 1 | 0,033 | 4 | 0,11 | P | Z |
Erythroxylum argentinum O. E. Schulz | 1,5 | 1 | 0,029 | 4 | 0,10 | Sí | Z |
Ilex dumosa Reissek | 7,5 | 1 | 0,031 | 4 | 0,10 | P | Z |
Ocotea silvestris Vattimo | 5,0 | 1 | 0,029 | 4 | 0,10 | Sí | Z |
Zanthoxylum astrigerum (R. S. Cowan) P. G. Waterman | 5,0 | 1 | 0,027 | 4 | 0,10 | Sí | Au |
Diospyros inconstans Jacq. | 6,0 | 1 | 0,015 | 4 | 0,09 | San | Z |
Eugenia uniflora L. | 4,0 | 1 | 0,018 | 4 | 0,09 | C | Z |
Banara parviflora (A. Gray) Benth. | 2,5 | 1 | 0,005 | 4 | 0,08 | Sí | Z |
Cedrela fissilis Vell. | 2,5 | 1 | 0,006 | 4 | 0,08 | San | Au |
Ilex pseudobuxus Reissek | 4,0 | 1 | 0,003 | 4 | 0,08 | P | Z |
Machaerium stipitatum (DC.) Vog | 3,5 | 1 | 0,005 | 4 | 0,08 | P | En |
Myrcia tijucensis Kiaersk. | 3,0 | 1 | 0,006 | 4 | 0,08 | C | Z |
Psychotria brachyceras Müll. Arg. | 2,0 | 1 | 0,003 | 4 | 0,08 | San | Z |
Vitex megapotamica (Spreng.) Moldenke | 2,0 | 1 | 0,004 | 4 | 0,08 | Sí | Z |
Campomanesia xanthocarpa O. Berg | 2,5 | 1 | 0,002 | 4 | 0,08 | P | Z |
Ouratea salicifolia (A. St. Hill. et Tul.) Engl. | 3,5 | 1 | 0,002 | 4 | 0,08 | San | Z |
Total | | 690 | 41,612 | 4.240 | 100 | | |
1
Revista Ouricuri, Juazeiro, Bahia, v.14, n.2. 2024, p.03 - 21. Jul./dez., Publicação contínua http://www.revistas.uneb.br/index.php/ouricuri | ISSN 2317-0131
Analizando la comunidad arbustiva en términos de grupos ecológicos, de las 81 especies encontradas, el 23% eran pioneras, el 26% secundarias tempranas, el 31% secundarias tardías y el 20% clímax. En cuanto a las estrategias de dispersión, de las 81 especies muestreadas, 74 (91%) presentaban dispersión zoocórica (Tabla 1).
En la regeneración natural de especies arbustivas y arbóreas (Tabla 2), se encontraron 67 especies de 30 familias botánicas, considerando las dos clases tamaño. De ellas, el 31% se registraron en la clase 1 (plántulas) y el 69% en la clase 2 (juveniles). Las familias más representativas, combinando las dos clases de tamaño, fueron Myrtaceae con 13 especies, totalizando el 16% de RNT; y Lauraceae con siete. Algunas especies sólo aparecieron en el componente de regeneración, como Daphnopsis racemosa, Euterpe edulis, Guarea macrophylla, Luehea divaricata, Myrcia anacardiifolia, Psychotria sp., P. suterella y Zollernia ilicifolia. Sorocea bonplandii presentó el valor más alto de regeneración natural (13,5%), seguida de Esenbeckia grandiflora (7,8%), Nectandra oppositifolia (7,7%), Sapium glandulosum (6,7%) y Eugenia brevistyla (3,4%). El valor más bajo (0,2%) se estimó para 15 especies, presentes únicamente en la clase de altura 2 (juveniles).
El análisis de correspondencias, aplicado a la matriz que contenía los datos de los estudios realizados en el sur de Brasil, indicó que las zonas estaban separadas a lo largo de los dos primeros ejes de ordenación (Figura 3). Aunque se considera una técnica descriptiva y exploratoria, el Análisis de Correspondencias simplifica los datos complejos y produce análisis de información que apoyan las conclusiones. La separación de las áreas indica que el sitio de este estudio parece ser una zona de contacto entre las formaciones de restinga de Rio Grande do Sul y las áreas de bosque denso ombrofilo de Rio Grande do Sul y Santa Catarina.
En cuanto a la distribución geográfica y los corredores migratorios, la mayoría de las especies se encontraron ampliamente distribuidas, especialmente S. bonplandii y G. concolor, que contribuyeron significativamente a la densidad de individuos en el área de estudio. Entre las especies del contingente atlántico, destacaron las siguientes: Coussapoa microcarpa, Erythroylum argentinum, Esenbeckia grandiflora, Eugenia schuechiana, Faramea marginata, Ficus organensis, Garcinia gardneriana, Guapira opposita, Myrcia glabra, Myrcia multiflora y Nectandra oppositifolia.
3. DEBATE
Según los datos disponibles en el Plan de Manejo del PEVA, se conocen 806 especies de plantas vasculares en la unidad de conservación. Aunque este entre las unidades de conservación con la vegetación.
Tabla 2 - Regeneración natural de especies arbustivas de restinga en el Parque Estadual de Itapeva, Torres, Rio Grande do Sul.
Table 2 - The shrub-trees species natural regeneration in a Restinga at Parque Estadual de Itapeva, Torres Municipality, Rio Grande do Sul, Brazil.
Tabela 2 - The shrub-trees species natural regeneration in a Restinga at Parque Estadual de Itapeva, Torres Municipality, Rio Grande do Sul, Brazil
Especies | RNC1 | RNC2 | RNT |
Sorocea bonplandii | 4,82 | 8,72 | 13,54 |
Esenbeckia grandiflora | 1,54 | 6,25 | 7,79 |
Nectandra oppositifolia Sapium glandulosum | 4,01 4,36 | 3,68 2,34 | 7,69 6,7 |
Eugenia neoaustralis | 0 | 3,41 | 3,41 |
Marlierea eugeniopsoides | 1,34 | 1,94 | 3,28 |
Cupania vernalis | 0,6 | 2,67 | 3,27 |
Mollinedia schottiana Myrcia pubipetala | 2,89 0,47 | 0 2,41 | 2,89 2,87 |
Psychotria brachyceras | 0,27 | 2,35 | 2,61 |
Eugenia schuechiana | 0,2 | 2,34 | 2,54 |
Myrsine umbellata | 1,54 | 0,8 | 2,34 |
Persea willdenovii | 0 | 2,21 | 2,21 |
Maytenus sp. | 0 | 2 | 2 |
Aiouea saligna | 0,6 | 1,27 | 1,87 |
Guapira opposita | 0,2 | 1,47 | 1,67 |
Alchornea triplinervia | 0,4 | 1,21 | 1,61 |
Posoqueria latifolia | 0,6 | 1 | 1,6 |
Gymnanthes concolor | 0,4 | 1,14 | 1,54 |
Psidium cattleianum | 0,47 | 1,07 | 1,54 |
Daphnopsis racemosa | 0 | 1,47 | 1,47 |
Endlicheria paniculata | 0,6 | 0,8 | 1,4 |
Trichilia lepidota | 0,6 | 0,74 | 1,34 |
Allophylus edulis | 0,2 | 1,14 | 1,34 |
Prunus myrtifolia | 0 | 1,27 | 1,27 |
Solanum pseudoquina | 0 | 1,27 | 1,27 |
Nectandra lanceolata | 0,2 | 0,94 | 1,14 |
Faramea montevidensis | 0,4 | 0,74 | 1,14 |
Chrysophyllum inornatum | 0,2 | 0,87 | 1,07 |
Myrsine coriacea | 0 | 1,07 | 1,07 |
Ocotea pulchella | 0,4 | 0,47 | 0,87 |
Lithraea brasiliensis | 0 | 0,8 | 0,8 |
Myrcia brasiliensis | 0,4 | 0,4 | 0,8 |
Persea venosa | 0,4 | 0,4 | 0,8 |
Hirtella hebeclada | 0,2 | 0,4 | 0,6 |
Machaerium stipitatum | 0 | 0,6 | 0,6 |
Miconia sellowiana | 0,2 | 0,4 | 0,6 |
Schinus terebinthifolius | 0,2 | 0,4 | 0,6 |
Zanthoxylum rhoifolium | 0 | 0,6 | 0,6 |
Guarea macrophylla | 0,54 | 0 | 0,54 |
Symphyopappus casarettoi | 0 | 0,54 | 0,54 |
Erythroxylum cuspidifolium | 0 | 0,47 | 0,47 |
Erythroxylum deciduum | 0 | 0,47 | 0,47 |
Matayba guianensis | 0 | 0,4 | 0,4 |
Myrcia anacardiifolia Piper gaudichaudianum | 0,2 0,2 | 0,2 0,2 | 0,4 0,4 |
Ilex theezans | 0 | 0,34 | 0,34 |
Euterpe edulis Luehea divaricata | 0 0 | 0,27 0,27 | 0,27 0,27 |
Myrcia multiflora | 0,27 | 0 | 0,27 |
Psychotria suterella | 0,27 | 0 | 0,27 |
Blepharocalyx salicifolius | 0 | 0,2 | 0,2 |
Cabralea canjerana | 0 | 0,2 | 0,2 |
Casearia decandra | 0 | 0,2 | 0,2 |
Eugenia bacopari | 0 | 0,2 | 0,2 |
Eugenia uniflora | 0 | 0,2 | 0,2 |
Garcinia gardneriana | 0 | 0,2 | 0,2 |
Ilex pseudobuxus | 0 | 0,2 | 0,2 |
Mollinedia sp. | 0 | 0,2 | 0,2 |
Myrcia glabra | 0 | 0,2 | 0,2 |
Myrsine lorentziana | 0 | 0,2 | 0,2 |
Ouratea parviflora | 0 | 0,2 | 0,2 |
Psychotria sp. | 0 | 0,2 | 0,2 |
Tetrorchidium rubrivenium | 0 | 0,2 | 0,2 |
Vitex megapotamica | 0 | 0,2 | 0,2 |
Zollernia ilicifolia | 0 | 0,2 | 0,2 |
Total | 30,85 | 69,15 | 100 |
Esta afirmación fue corroborada en este estudio. En PEVA, al menos 67 especies de plantas vasculares están amenazadas de extinción y/o protegidas por ley en Rio Grande do Sul (RIO GRANDE DO SUL, 2003). De estas, tres especies fueron encontradas en el remanente estudiado (Euterpe edulis, Trichilia lepidota y Ocotea silvestris).
El número de especies encontradas en PEVA está por encima del estándar de riqueza esperado para los bosques arenosos de restinga en Rio Grande do Sul (Waechter, 1985; Dillenburg et al., 1992; Rossoni; Baptista, 1995; Avión; Mondin, 2001; Moretto; Mondin, 2002; Día; Waechter, 2004; Scherer et al., 2007). La representatividad de Myrtaceae, así como de los géneros Myrcia y Eugenia, sigue los patrones de riqueza para el bioma de la Mata Atlántica, sugiriendo afinidad del área de estudio con formaciones de este bioma (Sacramento et al., 2007; Murray-Smith et al., 2008).
La afinidad florística del área de estudio (Restinga) fue observada con estudios realizados en el Bosque Ombrófilo Denso en el sur de Brasil, principalmente en el sur de Santa Catarina (Figura 3). En este estudio, también se observó que no hubo especies de Sebastiania descritas entre las más importantes en estudios en las restingas de Rio Grande do Sul, donde, en muchos casos, predomina el componente arbustivo-arbóreo (Dillenburg et al., 1992; Rossoni; Baptista, 1995; Avión; Mondin, 2001; Día; Waechter, 2004; Scherer et al., 2005). A diferencia de los estudios citados, en este estudio los valores de densidad más altos se atribuyeron a S. bonplandii, M. umbellata y C. inornatum. M. umbellata tampoco fue mencionada entre las más abundantes en los bancos de arena del estado. Sorocea bonplandii se destacó como una de las especies más importantes en los estudios realizados en bosques estacionales (Jarenkow; Waechter, 2001; Giehl; Jarenkow, 2008) y ombrófilos densos (Citadini-Zanette et al., 2003).
De acuerdo con los estudios desarrollados por Rambo (1950), la mayoría de las especies tropicales originarias del norte y centro de Brasil migraron al Rio Grande desde el Sur, a través de la "Porta de Torres", la Mata Atlántica es una vía de comunicación entre el sur de Santa Catarina y el nordeste de Rio Grande do Sul. La mayoría de las especies del contingente atlántico ocupan el sotobosque del bosque.
Los elevados valores de densidad obtenidos por especies de amplia distribución (por ejemplo, S. bonplandii) reflejan la gran plasticidad ecológica de estas especies y su capacidad para adaptarse a entornos estresantes. La colonización de un nuevo entorno suele producirse a partir de tasas reproductivas positivas y condiciones ambientales similares, proporcionando la capacidad de competencia interespecífica que conduce a la permanencia y desarrollo de la nueva especie en la zona (PEARMAN et al., 2008). Las tasas reproductivas positivas suelen estar relacionadas conlainteraccióncon agentes polinizadores y dispersores. La dispersión es una importante interacción entre fauna y flora, constituyendo uno de los mecanismos esenciales para la dinámica forestal y, consecuentemente, influyendo en la regeneración natural de las poblaciones (Zambonim, 2001; Tabarelli; Peres, 2002).
En este estudio, la zoocoria representó el 91% del número total de especies muestreadas. Este patrón también fue identificado por Scherer et al. (2005) en un componente arbóreo de restinga arenosa en el Parque Estatal de Itapuã, RS. Estos estudios confirmaron la importancia de los agentes bióticos en el flujo genético en las restingas arenosas, similar a los resultados de varios autores, como el modo más importante de dispersión de especies leñosas en la Mata Atlántica.
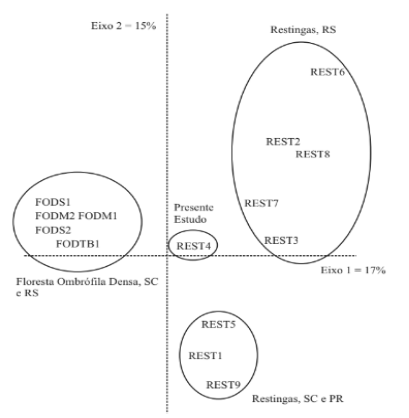
Figura 3 – Análisis de correspondencia de las formaciones de Mata Atlántica en el sur de Brasil listadas en la metodología, donde: FODM - Bosque Ombrófilo Denso de Montaña; FODS - Bosque Ombrófilo Denso Submontano; FODTB - Bosque Ombrófilo Denso de Tierras Bajas, REST Restinga.
Figure 3 – Atlantic Forest formations correspondence analyses in the South of Brazil listed in the methodology, where: FODM - Mountain Dense Ombrophilous Forest; FODS - Submountain Dense Ombrophilous Forest; FODTB - Lowlands Dense Ombrophilous Forest, REST Restinga.
Figura 3 – Análise de correspondência das formações da Mata Atlântica no Sul do Brasil listadas na metodologia, onde: FODM - Floresta Ombrófila Densa Montanhosa; FODS - Floresta Ombrófila Densa Submontanhosa; FODTB - Floresta Ombrófila Densa de Terras Baixas, REST Restinga.
El patrón de distribución geográfica de los estudios, obtenido mediante análisis de correspondencias, remite a la proximidad geográfica como causa principal de mayor similitud entre áreas (patrón geográfico), a menudo corroborada por condiciones ambientales similares (clima, temperatura, precipitaciones, suelo, etc.), como se observa en las formaciones de Bosque Denso Ombrofilo Atlántico. Sin embargo, áreas geográficamente distantes con condiciones ambientales similares también pueden mostrar similitudes en su composición (Guarino; Walter, 2005). Estos patrones pueden influir en la clasificación de la vegetación, unas veces basada en aspectos florísticos (especies dominantes y frecuentes) y otras en aspectos ambientales (Lima et al., 2009).
Analizando los resultados de la regeneración natural de especies arbustivas y arbóreas, se observó que las 10 primeras especies acumularon el 54% de la regeneración Comunidad estudiada oscilaron entre el 13,5% y el 0,2%. Scherer et al. (2005) obtuvieron tasas de regeneración natural total en un bosque de restinga arenosa que oscilaban entre el 22,2% y el 0,6%, superiores a los resultados de este estudio. La presencia restringida de algunas especies en el componente en regeneración puede indicar una afluencia de nuevas especies a la comunidad, o apuntar a la rareza de estas poblaciones en la comunidad, o incluso al hecho de que les resulta más difícil asentarse en la comunidad estudiada debido a sus condiciones ambientales. Euterpe edulis sólo fue muestreada en el componente regenerador, destacando el papel de la "Porta de Torres" como importante ruta migratoria de especies contingentes del Atlántico en Rio Grande do Sul. Esta especie es considerada un representante típico de este contingente florístico, avanzando hacia la porción norte del bosque en las laderas de la Serra do Sudeste y, a lo largo de las laderas sur de la Meseta Sur Brasileña, hacia la región central del estado (Jarenkow; Waechter, 2001).
Las menores tasas de regeneración estimadas en la zona de estudio pueden atribuirse a diversos factores, entre ellos el tipo y propiedades del suelo en el tramo analizado, ya que en Itapeva, ubicado en el municipio de Torres, predominan los suelos arenosos, con una mayor deposición de hojarasca en las zonas bajas entre las dunas. Un aspecto importante relacionado con la estrategia de regeneración es la densidad de adultos en la comunidad, así como la estrategia de crecimiento. Los árboles del dosel necesitan muchos más recursos para reproducirse que las especies del sotobosque, que normalmente se reproducen a la sombra y tienen una estructura morfológica más simple (Comita et al., 2007), como demuestra S. bonplandii.
En el caso de formaciones pioneras como las restingas, la mayoría de las especies encontradas están asociadas a su capacidad para tolerar el estrés provocado por el medio, lo que refleja una gran plasticidad ecológica por parte de las especies, que a menudo no encajan en los grupos ecológicos típicos aplicados a los bosques tropicales.
Relacionando aspectos de regeneración y grupos ecológicos, la tolerancia a la sombra puede establecerse como una estrategia importante para mantener la alta diversidad y coexistencia de especies, ya que son capaces de establecerse y permanecer durante mucho tiempo, en el sotobosque. En el otro extremo del espectro, las plantas dependientes de la luz necesitan "acontecimientos fortuitos" (Brokaw; Busing, 2000) como la apertura del dosel para sobrevivir, lo que conduce a un menor número de individuos en el banco de plántulas en comparación con las especies tolerantes a la sombra. Este patrón lleva a esperar una mayor coexistencia de especies en el sotobosque en detrimento del dosel. Distinguir las estrategias de coexistencia con respecto a las etapas del ciclo de vida puede revelar mecanismos que operan dentro y entre ambientes, proporcionando una base para dilucidar la compleja relación en las comunidades vegetales a lo largo de gradientes ambientales (Martins, 2010).
Las características ecológicas están cada vez más presentes en las hipótesis de coexistencia, evidenciadas por mecanismos importantes como intercambios obligatorios (trade off), formas de vida y etapas ontogenéticas del ciclo vital, configurando cuatro factores ambientales e históricos (1) heterogeneidad horizontal; (2) heterogeneidad vertical;
(3) régimen de perturbación; y (4) interacciones bióticas. Estos factores contribuyen a la coexistencia de especies entre las etapas de vida de las plantas (Nakashizuka, 2001).
4. CONCLUSIÓN
A través del análisis florístico-estructural y fitogeográfico realizado en la restinga y en los bosques ombrófilos del sur de Brasil, se puede afirmar que la restinga en cuestión presenta una elevada riqueza específica en comparación con otros relevamientos regionales, lo que probablemente se deba al predominio de elementos florísticos del contingente oriental y de especies ampliamente distribuidas. Debido al elevado número de especies presentes, la estructura de la comunidad y la regeneración natural encontrada, así como la contribución del Bosque Denso Ombrofilo Atlántico, se obtuvieron resultados diferentes a los de otros relevamientos realizados sobre la vegetación de la restinga en el sur de Brasil, destacando la importancia de la "Porta de Torres" como corredor de vegetación para Rio Grande do Sul.
En este sentido, este estudio ha añadido nueva información a nuestro conocimiento de la estructura comunitaria de la Restinga en Rio Grande do Sul. Los resultados aquí presentados contribuirán a nuevos estudios sobre la dinámica del PEVA y a programas de gestión, preservación y recuperación del medio ambiente.
REFERENCIAS
ARAÚJO, D. S. D.; LACERDA, L. D. A naturaleza de los bancos de arena. Ciência Hoje, v.6, n.33, p.42-48, 1987.
ASSUMPÇÃO, J.; NASCIMENTO, M. T. Estructura y composición florística de cuatro formaciones de vegetación de restinga en el complejo lagunar de Grussaí/Iquipari, São João da Barra, RJ, Brasil. Acta Botanica Brasilica, v.14, n.3, p.301-315, 2000.
BROKAW, N.; BUSING, R. T. Niche versus chance and tree diversity in forest gaps. Trends in Ecology & Evolution, v.15, p.183-187, 2000.
CITADINI-ZANETTE, V. Florística, fitosociología aspectos de dinámica de un remanente forestal Atlántico en la cuenca del Río Novo, Orleans, SC. 1995. 236f. Tesis (Doctorado en Ecología) - Universidad Federal de São Carlos, São Carlos, 1995.
CITADINI-ZANETTE, V.; SANTOS, R.; SOBRAL, M. Estudio florístico de la vegetación arbustiva y arbórea en un área ecotonal entre restinga y bosque ombrofilo denso de tierras bajas (Praia de Palmas, Governador Celso Ramos, Santa Catarina, Brasil). Revista Tecnologia e Ambiente, v.7, n.1, p.105-120, 2001.
CITADINI-ZANETTE, V. et al. Mirtáceas del sur de Santa Catarina: un subsidio para la recuperación de ecosistemas degradados. Revista de Tecnologia e Ambiente, v.9, n.2, p.61-75, 2003.
COMITA, L. S. et al. Patterns of woody plant species abundance and diversity in the seedling layer of a tropical forest. Journal of Vegetation Science, v.18, p.163-174, 2007.
DILLENBURG, L. R.; WAECHTER, J. L.; PORTO, M. L. Species composition and structure of a sandy coastal plain forest in northern Rio Grande do Sul, Brazil. En: SEELIGER, U. (Org.) Coastal plant communities communities of Latin America. San Diego: Academic Press, 1992.
DORNELES, L. P. P.; WAECHTER, J. L. Estructura del componente arbóreo del bosque de restinga arenosa del Parque Nacional Lagoa do Peixe, Rio Grande do Sul. Hoehnea, v.31, n.1, p.61-71, 2004.
FALKENBERG, D. B. Aspecto da flora e da vegetação secundária da Restinga de Santa Catarina, sul do Brasil. Insula, n. 28, p.1-30, 1999.
FERRETTI, A. R. et al. Clasificación de especies arbóreas en grupos ecológicos para revegetación con nativas en el Estado de São Paulo. Florestar Estatístico, v.3, n.7, p.73-77, 1995.
FELFILI, J. M. et al. Procedimientos y métodos de muestreo de la vegetación. En: FELFILI, J. M. et al. (Org.) Fitosociología en Brasil: métodos y estudios de caso. Viçosa, MG: Universidade Federal de Viçosa, 2011. p.86-121.
FINOL, U. H. Nuevos parámetros a considerar en el análisis estructural de las selvas vírgenes
tropicales. Revista Forestal Venezolana, v.14, n.21, p.29-42, 1971.
FORZZA, R. C. et al. (Org.) Catálogo de plantas y hongos de Brasil. Río de Janeiro: Instituto de Investigación del Jardín Botánico de Río de Janeiro, 2010. 1699 p.
FREIRE, M. S. B. Levantamiento florístico del Parque Estatal de las Dunas de Natal. Acta Botanica Brasilica, v. 4, n. 2, p. 41-59, 1990.
GIEHL, E. L. H.; JARENKOW, J. A. Gradiente en el componente arbóreo y relación con la inundación en un bosque ribereño, Río Uruguay, sur de Brasil. Acta Botanica Brasilica, v.22, n.3, p.741-753, 2008.
GUARINO, E. S. G.; WALTER, B. M. T. Fitosociología de dos tramos inundados de bosque de galería en el Distrito Federal, Brasil. Acta Botanica Brasilica, v.19, n.3, p.431-442, 2005.
HENSTSCHEL, R. L. Gradiente de vegetación, variables ambientales y restauración en el arenal de Praia do Ouvidor, Garopaba, Santa Catarina. 2008. 76f. Disertación (Maestría en Botánica) - Universidad Federal de Rio Grande do Sul, Porto Alegre, 2008.
INSTITUTO NACIONAL DE METEOROLOGÍA, MINISTERIO DE AGRICULTURA, GANADERÍA Y PESCA SUMINISTRO - INMET. Disponible en:
<http://www.inmet.gov.br>. Fecha de consulta: 28 de octubre de 2009.
JARENKOW, J. A. Estudio fitosociológico comparativo entre dos áreas de bosque de ladera en Rio Grande do Sul. 1994. 122f. Tesis (Doctorado en Ecología y Recursos Naturales) - Universidad Federal de São Carlos, São Carlos, 1994.
JARENKOW, J. A.; WAECHTER, J. L. Composición, estructura y relaciones florísticas del componente arbóreo de un bosque estacional en Rio Grande do Sul, Brasil. Revista Brasileira de Botânica v.24, n.3, p.263-272, 2001.
KENT, M.; COKER, P. Descripción y análisis de la vegetación. Nueva York: Wiley, 1992. 363 p.
LIMA, J. R. et al. Floristic composition of the Montane Seasonal Deciduous Forest of Serra das Almas, CE, Brazil. Acta Botanica Brasilica, v.23, n.3, p.756-763, 2009.
LINDEMAN, C. A. M. Vegetación en Rio Grande do Sul. Porto Alegre: Universal, 1906. 365 p.
MAGNAGO, L. F. S.; MARTINS, S. V.; PEREIRA, O. J. Heterogeneidad florística de las fitocenosis de restinga en los estados de Río de Janeiro y Espírito Santo, Brasil. Revista Árvore, v.35, n.2, p.245-254, 2011.
MAGNAGO, L. F. S. et al. Phytophysiognomic-edaphic gradient in Restinga forest formations in southeastern Brazil. Acta Botanica Brasilica, v.24, n.3, p.734-746, 2010.
MARTINS, R. Florística, estructura fitosociológica e interaciones interespecíficas de aremanente de Ombrofilo Denso Bosque como subsidio para la recuperación de zonas degradadas por la minería minería Siderópolis, SC. 2005. 93f. Disertación (Biología Vegetal) - Universidad Federal de Santa Catarina, Florianópolis, 2005.
MARTINS, R. Composición y estructura de la vegetación en diferentes formaciones en la selva atlántica, sur de Santa Catarina, Brasil. 2010. 148f. Tesis (Doctorado en Botánica)- Universidad Federal de Rio Grande do Sul, Porto Alegre, 2010.
MORAES, D.; MONDIN, C. A. Florística y fitosociología del estrato arbóreo del bosque arenoso de Balneário Quintão, Palmares do Sul/ RS. Pesquisas, Série Botânica, n.51, p.87-100, 2001.
MORETTO, F.; MONDIN C. A. Pesquisa quali- cuantitativa do componente leñoso do sotobosque de uma floresta de restinga arenosa em Balneário Quintão, Palmares do Sul/RS, Porto Alegre. Pesquisas, Série Botânica, n.52, p.111-123, 2002.
MUELLER-DOMBOIS, D.; ELLENBERG, H. Objetivos y métodos de ecología de la vegetación. Nueva York: Wiley, 1974. 574 p.
MURRAY-SMITH, C. et al. Plant diversity hotspots in the atlantic coastal forests of Brazil. Conservation Biology, v.23, n.1, p.151-163, 2008.
NAKASHIZUKA, T. Species coexistence in temperate, mixed deciduous forests. Trends in Ecology & Evolution, v.16, n.4, p.205-210, 2001.
NEGRELLE, R. R. B. Composición florística y estructura vertical de un tramo de Bosque Ombrofilo Denso en la Llanura Cuaternaria. Hoehnea, v.33, n.3, p.261-289, 2006.
PEARMAN, P. B. et al. Niche dynamics in space and time. Trends in Ecology & Evolution, v.23, n.3, p.149-158, 2008.
PORTO ALEGRE. Secretaría de Estado de Medio Ambiente de Rio Grande do Sul - SEMA. Plan de manejo del Parque Estadual de Itapeva. Porto Alegre: 2006. 259 p.
RAMBO, B. La puerta de las torres. Anales botánicos del Herbário Barbosa Rodrigues, n.2, p.9-20, 1950.
RIO GRANDE DO SUL. Decreto nº ?42099 de 31 de diciembre de 2002. Lista de especies de flora nativa amenazadas de extinción en Rio Grande do Sul. Boletín Oficial del Estado de Rio Grande do Sul, p.1-6, 2003.
ROCHA, C. F. D. et al. Tetrápodos endémicos y amenazados en las restingas de los corredores de biodiversidad de Serra do Mar y de la Mata Atlántica central en Oriente. Revista Brasileira de Biologia, v.65, n.1, p.159-168, 2005.
ROSSONI, M. G.; BAPTISTA, L. R. M. Composición florística del bosque de restinga, Balneário Rondinha Velha, Arroio do Sal, RS, Brasil. Investigación, Serie Botánica, n.45, p.115-131, 1995.
SACRAMENTO, A. C.; ZICKEL, C. S.; ALMEIDA JUNIOR, E. B. Aspectos florísticos de restinga no litoral de Pernambuco. Revista Árvore, v.31, n.6, p.1121-1130, 2007.
SCHERER, A.; MARASCHIN-SILVA, F.; BAPTISTA, L. R. M. Regeneración arbórea en un pastizal de restinga en Rio Grande do Sul, Brasil. Iheringia , Série Botânica , v.62, n.1-2, p.89-98, 2007.
SCHERER, A.; SILVA, F. M.; BAPTISTA, L. R. M. Florística y estructura del componente arbóreo de los bosques arenosos de restinga en el Parque de Itapuã, RS, Brasil. Acta Botanica Brasilica, v.19, n.4, p.717-726, 2005.
SILVA, S. M. Composición florística y fitosociología de un trecho de bosque de restinga en Ilha do Mel, Paranaguá, PR. 1990. 146f. Disertação (Maestria em Biologia Vegetal) - Universidade Estadual de Campinas, São Paulo, 1990.
SILVA, S. M.; BRITEZ, R. M. La vegetación de la Llanura Costera. En: MARQUES, M. C. M.; BRITEZ, R. M. (Org.). História natural e conservação da Ilha do Mel. Curitiba: Universidad Federal de Paraná, 2005. p.49-84.
TABARELLI, M.; PERES, C. A. Abiotic and vertebrate seed dispersal in the Brazilian Atlantic Forest: implications for forest regeneration. Biological Conservation, v.106, p.165-176, 2002.
van der PIJL, L. Principios de dispersión en plantas superiores. 2.ed. Berlín: Springer, 1972. 215 p.
VOLPATO, M. M. L. Regeneración natural en un bosque secundario del dominio forestal atlántico: un análisis fitosociológico. 1994. 123f. Disertação (Maestria) - Universidade Federal de Viçosa, Viçosa, MG, 1994.
WAECHTER, J. L. Aspectos ecológicos da vegetação de restinga no Rio Grande do Sul, Brasil. Comunicaciones del Museo de Ciencias de la PUCRS, Serie Botánica, n.33, p.49-68, 1985.
ZAMBONIM, R. M. Base de datos como subsidio para la conservación y restauración en tipologías de vegetación del Estado Parque Estadual da Serra do Tabuleiro. 2001. 118f. Disertación (Maestría en Recursos Fitogenéticos) - Universidad Federal de Santa Catarina, Florianópolis, 2001.
1
Revista Ouricuri, Juazeiro, Bahia, v.14, n.2. 2024, p.03 - 21. Jul./dez., Publicação contínua http://www.revistas.uneb.br/index.php/ouricuri | ISSN 2317-0131